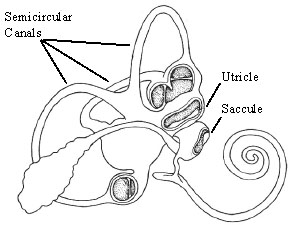
The vestibular senses are made possible by the vestibular labyrinth, found within the inner ear. The entire labyrinth is about the size of a marble and is located next to the cochlea, used in hearing.
Figure 1. The Vestibular Labyrinth, showing the semicircular canals and the otolith organs. (Freidmann and Ballantyne, 1984)
The vestibular labyrinth consists of five parts: three semicircular canals, and the two otolith organs, known as the utricle and the saccule. All of these sections are connected by bony passageways filled with a endolymphatic fluid. The labyrinth also connects to the cochlea, which is responsible for hearing.
The canal consists of a circular tube (much like the inner-tube of a bicycle) filled with viscous endolymphatic fluid. At one end of the tube is a small triangular shaped mass of gelatinous material called the cupula, which partially blocks the canal. The cupula sits on a bulged area covered with hair cells, known as the crista.
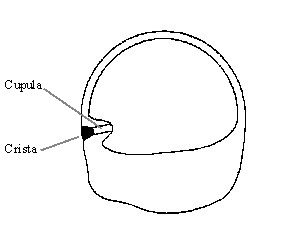
Figure 2. A cross-section of a semicircular canal. (Grant, 1995)
When the head is rotated, the fluid lags behind because of inertia. The fluid pushes against the cupula, deflecting it. This deflection is measured by the hair cells in the crista.
Each otolith organ consists of a cavity in the bony labyrinth. Within this cavity lies a flat mass of small dense bodies called otoconia or otoliths (literally: ear stones). These are held together by an extracellular matrix. This mass, called the otoconial layer, is connected to the wall of the labyrinth on one side by a viscoelastic gel, conveniently called the gel layer. The labyrinth wall where the gel layer attaches is full of hair cells. The entire cavity is filled with a viscous fluid.
When the head is moved, the dense mass initially lags behind due to inertia. This causes the gel layer to shear, which is sensed by the hair cells beneath. Under the influence of gravity, the denser otoconial layer will deflect towards the earth, causing shear also.
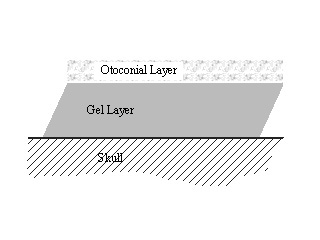
Figure 3. A schematic of the cross section of the otolith organ.
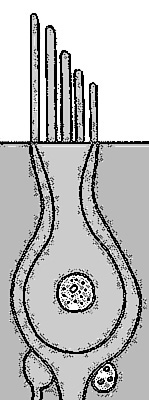
Figure 4. A diagram of a hair cell, showing the apical surface and cilia extending from it. (adapted from Freidmann and Ballantyne, 1984)
The top of the hair cell consists of an epithelial surface, a number of stereocilia, and a single kinocilium. There may be anywhere from ten to one hundred stereocila in a bundle. The kinocilium is thicker and, usually, taller than the stereocilia. Hair cells are packed in arrays where each cilia has six neighbors an equal distance apart (hexagonal packing).
The stereocilia are interconnected by small fibers called subapical bands. These bands extend down the length of the cilia and connect the cilia to each of its neighbors. Each stereocilia also connects to its next tallest neighbor by a single tip link. It is thought that tension in these tip links caused by the bundle being deflected opens small channels in the cilia. As these channels open, ions from the surrounding fluid rush in due to an electrochemical gradient, causing the cell to fire.
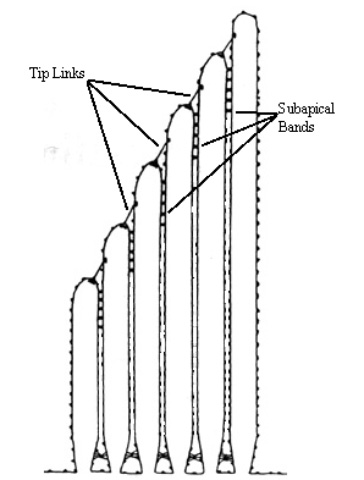
Figure 5. Schematic of hair cell showing interconnections. (adapted from Goodyear and Richardson, 1994)
Hair cells vary greatly from species to species, organ to organ, and even the location within the same organ. These variations lead us to ask many questions such as:
How do variations in bundle dimensions effect hair bundle stiffness?
What effect does stiffness have on the filtering ability of a hair cell?
Why are certain types of hair cells found in different areas of the vestibule?
In a more general sense, we have other questions dealing with the actual hair cell mechanism. These questions include:
Does tip link tension account for the observed cell output?
What is the structure and material that makes up the subapical bands?
For most tall, slender structures under transverse loading, the deflection due to shear is negligible when compared to the deflection due to bending. However, stereocilia have been shown to exhibit significant shearing. This indicates we must use the more complex Timoshenko, or Shear Deformable Theory in our calculations, rather than the more common, and easier to use, Euler Theory.
Cilia interconnections are assumed to be thin strands of proteins. It is widely believed that these interconnections will only carry a tensile force and will give no resistance to compression. Hence the relationship between deflection and force is nonlinear for the interconnections. This results in the first of our nonlinearities.
The second of our nonlinearities is due to geometry. As the bundle deflects, the interconnections also move and become reoriented. This means, the line of action of their forces varies with deflection.
Because our problem is nonlinear, we must modify our analysis. Finite Element Analysis is, by itself, a linear technique. We solve this problem by breaking our problem up into many small linear steps. We apply our force to the stereocilia and determine the deflection of the stereocilia. We then use that deflection to determine the forces generated by the interconnections and apply those forces to the stereocilia as well.
Cilia positions and deflections are read from the output file and converted into Inventor format. The resulting file can be displayed in an Inventor viewer, which provides full rotation, zoom, and animation capabilities. Unfortunately, its exporting capabilities and portability are limited.
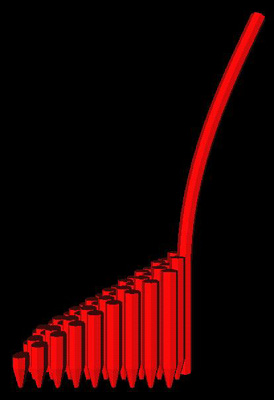
Figure 6. Output of Inventor format on SGI. A utricular hair cell is depicted.
To get around this, a number of tools were created using PV-Wave. All tools are run from a main driver routine. Different options are selected from a pull down menu. The tools enable the bundle to be viewed as a shaded surface or a simple line drawing. Both styles allow rotations, zooming, and scaling of deflections. The user can also animate the above. Below we have the line drawing of a bundle using the "line draw" feature. Deflections are scaled from zero to one, which gives the impression the bundle is being deflected. Further below is the full shaded bundle.
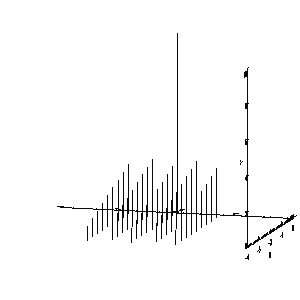
Figure 7. Line drawn output from PV-Wave

Figure 8. PV-Wave depiction of utricular hair cell.
Gaudin, A. J., and K. C. Jones, 1989. Human Anatomy and Physiology, Harcourt Brace and Jovanovich, Washington, D.C.
Grant, J.W., 1995. Chapter 39: Vestibular Mechanics, in The Bioengineering Handbook, CRC Press, Inc., Boca Raton, FL.
Furness, D. N. and C. N. Hackney, 1985. Crosslinks Between Stereocilia in the Guinea Pig Cochlea, Hearing Research, 18:177-188.
Goodyear, R. and G. Richardson, 1994. Differential Glycosylation of Auditory and Vestibular Hair Bundle Proteins Revealed by Peanut Agglutinin, The Journal of Comparative Neurology, 345:267-278.
Tilney, L. G., et. al., 1983. Actin Filaments, Stereocilia, and Hair Cells of the Bird Cochlea. II. Packing of Actin Filaments in the Stereocilia and in the Cuticular Plate and What Happens to the Organization When the Stereocilia Are Bent., Journal of Cell Biology, 96:822-834.
Duncan, K. and J. W. Grant, 1996. A finite Element Model of Inner Ear Hair Bundle Micromechanics, in press.
Reddy, J. N., 1993. An Introduction to the Finite Element Method, McGraw Hill, New York.
main.pro, shade.pro, lines.pro, set_scl.pro, set_size.pro, set_rot.pro, show.pro, animate.pro